- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- For Contributors
- Policies
- E-Submission

- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- Policies
- For Contributors
Articles
- Page Path
- HOME > J. Microbiol > Ahead of print > Article
-
Research article
Prophase roles of replication protein A in crossover formation and meiotic progression -
Rose M. Lee, Keun Pil Kim*
 , Jeong H. Joo*
, Jeong H. Joo* -
DOI: https://doi.org/10.71150/jm.2604001
Published online: June 18, 2026
Department of Life Sciences, Chung-Ang University, Seoul 06974, Republic of Korea
- *Correspondence Keun Pil Kim kpkim@cau.ac.kr Jeong H. Joo jkiiklovehot@cau.ac.kr
• Received: April 1, 2026 • Revised: April 22, 2026 • Accepted: April 27, 2026
© The Microbiological Society of Korea
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- 40 Views
- 3 Download
ABSTRACT
- Meiotic recombination is initiated by programmed DNA double-strand breaks (DSBs), which are subsequently processed to generate single-stranded DNA (ssDNA). Replication protein A (RPA), a heterotrimeric ssDNA-binding complex, plays essential roles in DNA replication, repair, and recombination; however, the specific functions of RPA in meiotic recombination progression and chromosome morphogenesis remain unclear. Here, we investigate the role of RPA in recombination and meiotic progression by conditionally depleting Rfa1, the large subunit of the RPA complex, using an auxin-inducible degron (AID) system in Saccharomyces cerevisiae. We show that Rfa1 depletion causes severe defects in meiotic recombination, including impaired DSB processing, defective chromosome axis assembly, compromised synaptonemal complex formation, and failure of ZMM-dependent crossover recombination. Notably, inhibition of Mek1 protein kinase activity, which bypasses the recombination checkpoint, does not rescue these defects in Rfa1-depleted cells. Together, these findings identify RPA as a key factor that stabilizes recombination intermediates and coordinates prophase I events with chromosome synapsis and crossover formation during meiosis.
Introduction
Materials and Methods
Results
Discussion
Acknowledgments
This work was supported by grants from the National Research Foundation of Korea, funded by the Ministry of Science, ICT & Future Planning (No. RS-2026-25474247).
Conflict of Interest
The authors declare no conflict of interest.
Supplementary Information
Fig. S1.
Fig. 1.Conditional Rfa1 degradation via auxin-inducible degron alters DNA damage sensitivity and meiotic progression. (A) Schematic representation of the RFA1–AID construct and the auxin-inducible degron (AID) system. Upon auxin (IAA) treatment, RFA1–AID is ubiquitinated by the SCFTIR1 complex and subsequently degraded by the proteasome. (B) DNA damage sensitivity assay of wild-type (WT) and RFA1–AID strains. The 10-fold serial dilutions (from 10-1 to 10-5) were spotted onto YPD plates containing the DNA-damaging agents: 0.03% MMS, 40 mM HU, 20 μM CuSO4, or 2 mM IAA. Plates were incubated at 30°C for 2 days. (C) Meiotic nuclear divisions in WT and RFA1–AID strains. Data are presented as the mean ± standard deviation (SD) (N > 200/time point; three independent biological replicates). (D) Protein expression levels of Rfa1 and Pgk1 during meiosis in the RFA1–AID strain. CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, followed by IAA (2 mM) treatment at 2.5 h to induce Rfa1 degradation. Proteins were detected using anti-Myc (for RFA1–AID–9Myc) and anti-Pgk1 antibodies. Unprocessed blot images are shown in Fig. S1B.
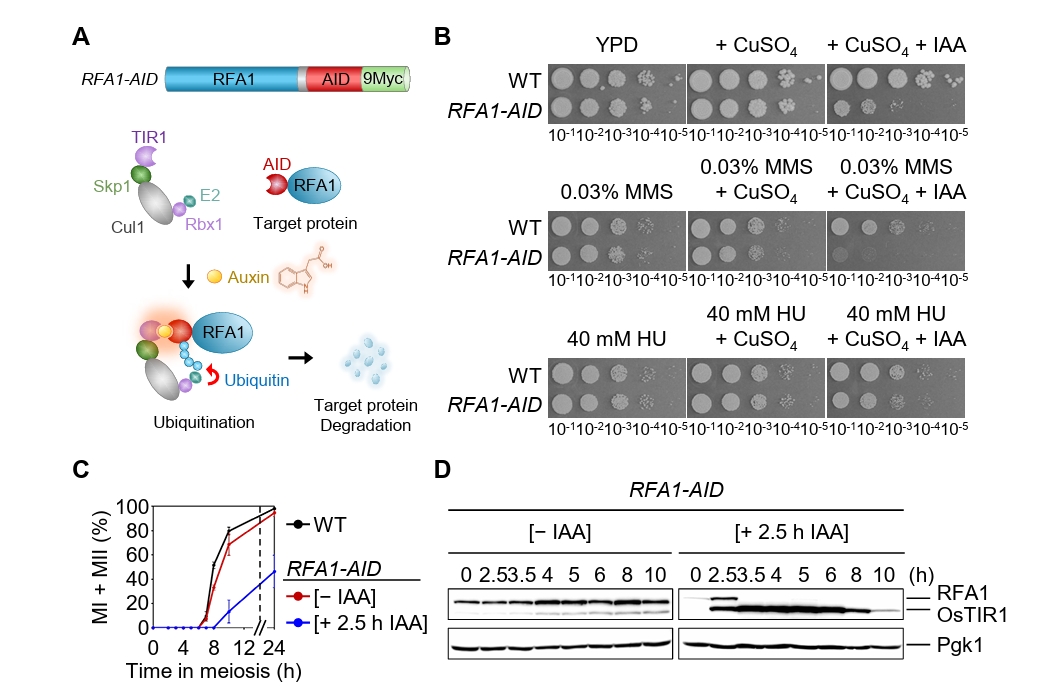
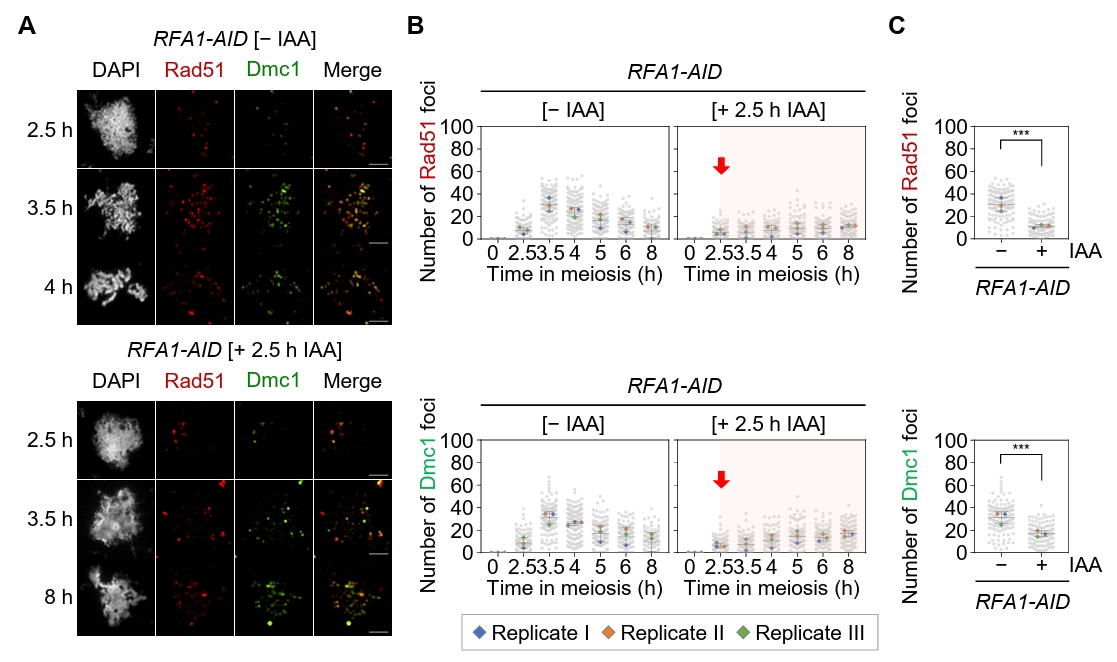
Fig. 2.Rfa1 is required for the formation of Rad51 and Dmc1 foci during meiotic recombination. (A) Representative immunofluorescence image of a meiotic nuclear spread from RFA1–AID strains in the absence [– IAA] or presence [+ IAA] of IAA. Cells were stained with anti-Rad51 (red) and anti-Dmc1 (green). CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to promote Rfa1 degradation. Scale bar = 2.5 μm. (B) Quantification of Rad51 and Dmc1 foci per nucleus in the RFA1–AID strain during meiotic recombination. Data are presented as the mean ± SD (three independent experiments). (C) Maximum peak time point of Rad51 and Dmc1 foci in the RFA1–AID strain shown in (B) [– IAA at 3.5 h] and [+ IAA at 8 h]. Data are presented as the mean ± SD (three independent experiments). P values for relevant comparisons are determined by Student’s t-test.
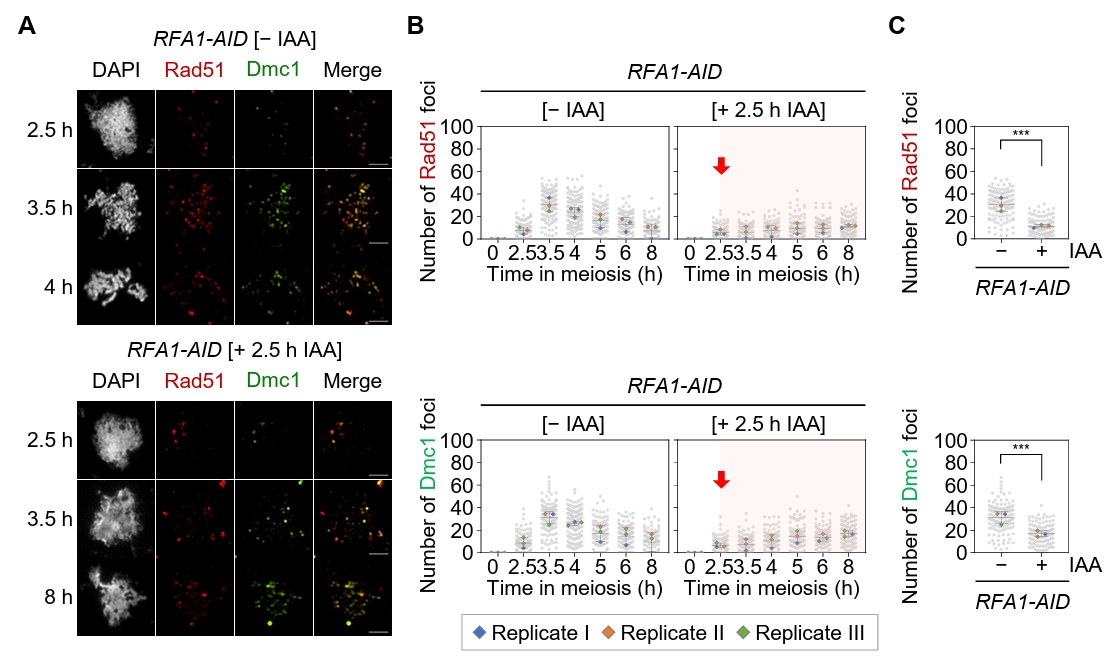
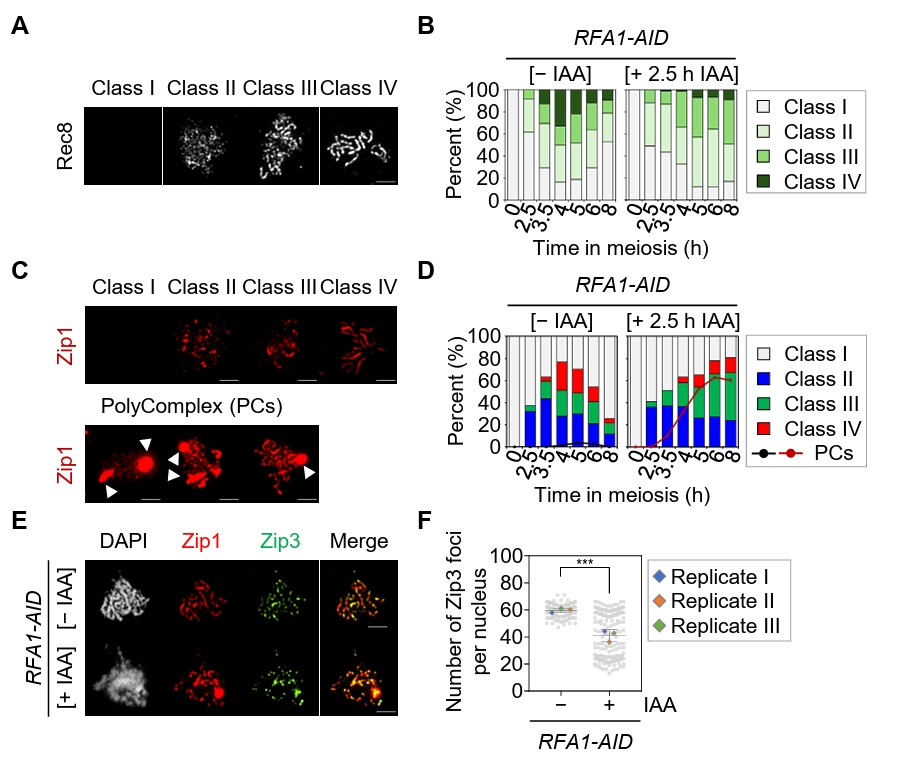
Fig. 3.Depletion of Rfa1 causes defects in meiotic recombination progression. (A) Representative immunofluorescence images of meiotic chromosome spreads stained with anti-Rec8 in the RFA1–AID strain. CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to induce Rfa1 degradation. Nuclear spreads were classified into four categories based on the Rec8 staining pattern: Class I, no staining; Class II, a modest number of chromosomes; Class III, extended or short linear chromosomes; Class IV, a full array of chromosomes. Scale bar = 2.5 μm. (B) Quantification of Rec8 staining classes at each time point shown in (A). (C) Representative images of meiotic chromosome spreads stained with anti-Zip1 in the RFA1–AID strain. CuSO4 and IAA were added as in (A). Nuclear spreads were classified into four categories based on the Zip1 staining pattern: Class I, blank; Class II, dotty chromosomes; Class III, short or discontinuous linear chromosomes; Class IV, full-array chromosomes. White arrowheads indicate polycomplexes (PCs). Scale bar = 2.5 μm. (D) Quantification of Zip1 staining classes at each time point shown in (C). The line graph indicates the percentage of cells with PCs. (E) Representative meiotic chromosome spreads from RFA1–AID strains immunostained with anti-Zip1 (red) and anti-Flag (for Zip3 staining, green). CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to induce Rfa1 degradation. Scale bar = 2.5 μm. (F) Quantification of Zip3 foci per nucleus at the peak time points of Zip3 expression during meiotic recombination: [– IAA] at 4 h and [+ IAA] at 6 h. Data are presented as the mean ± SD (three independent experiments). P values for relevant comparisons are determined by Student’s t-test.
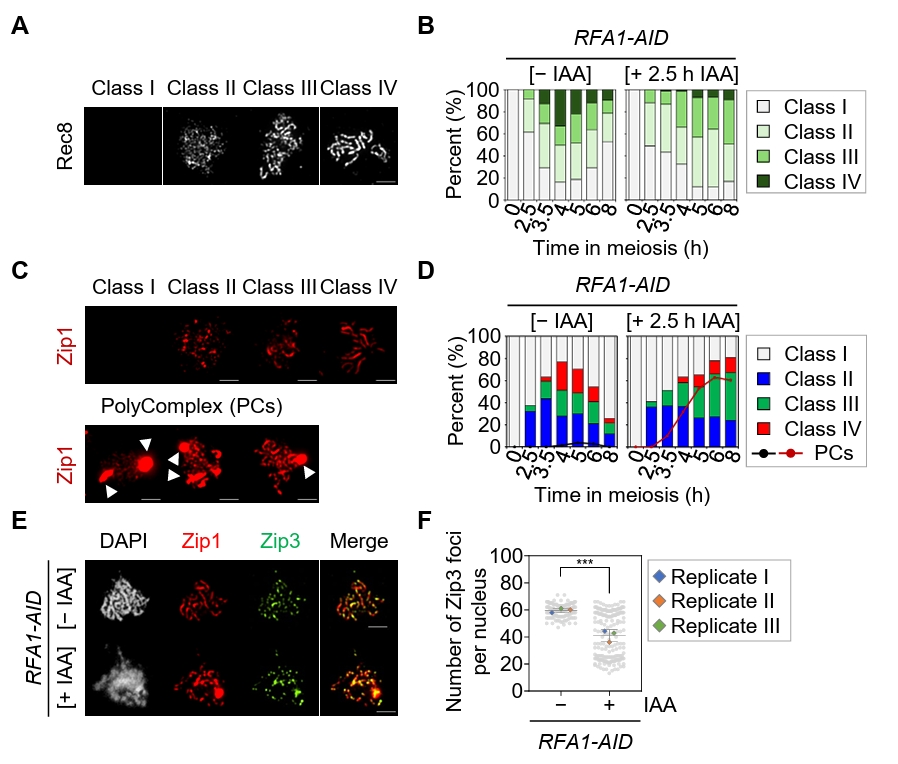
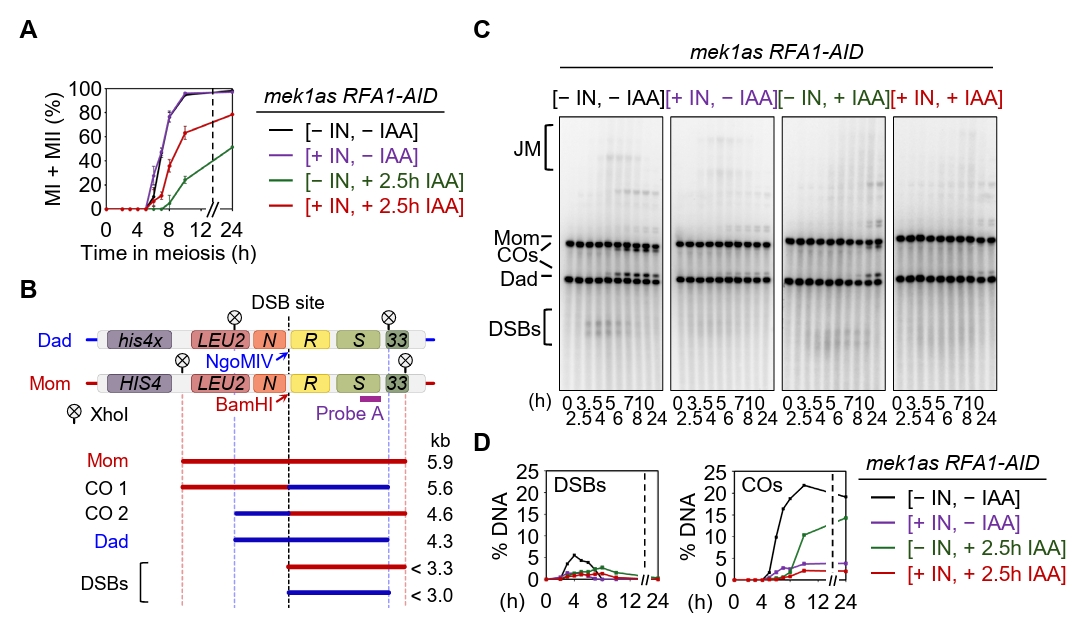
Fig. 4.Rfa1 is essential for meiotic DSB and crossover formation. (A) Meiotic nuclear division in the mek1as RFA1–AID strain. Mek1 kinase activity was inhibited by the addition of 1-NA-PP1 (IN; 2 µM) at 0 h. CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to induce degradation of Rfa1. Data are presented as the mean ± SD (N > 200 cells per time point; three independent biological replicates). (B) Schematic of the HIS4::LEU2 recombination hotspot, which generates double-strand breaks (DSBs) during meiosis. Positions of XhoI restriction sites are indicated by an X. Probe A (purple), used for Southern blot analysis, is shown. DSB, double-strand break; CO, crossover. (C) Representative one-dimensional gel images from mek1as RFA1–AID strains. (D) Quantification of DSB and CO levels shown in (C).
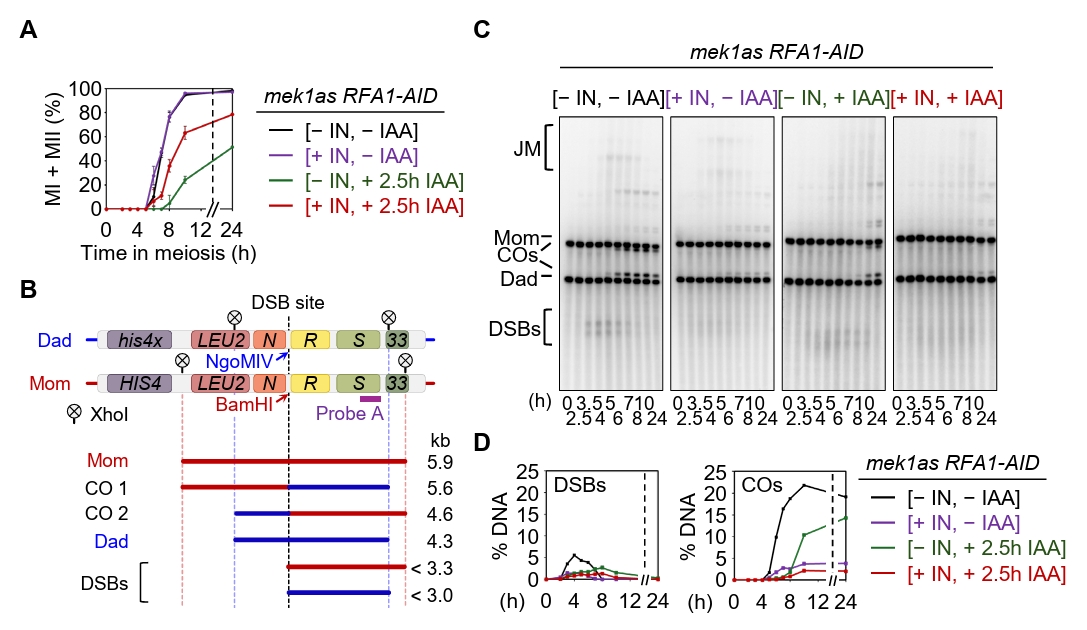
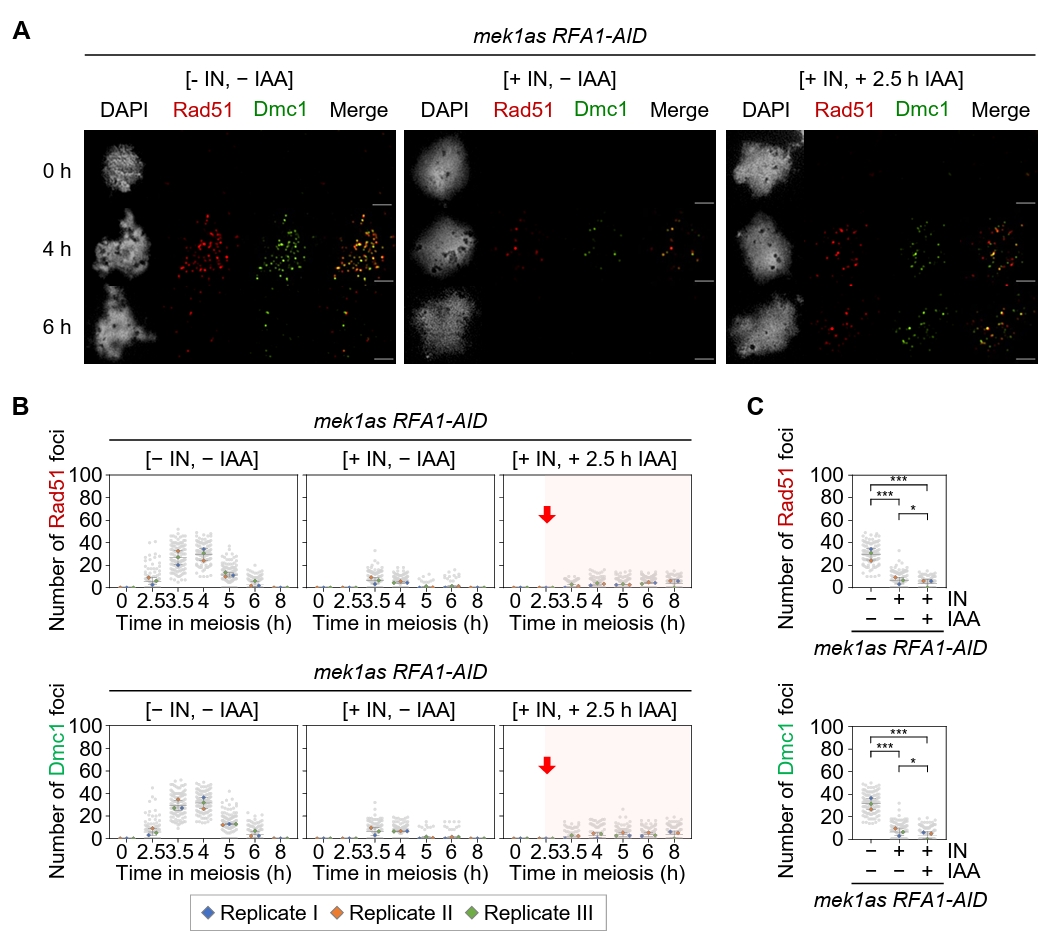
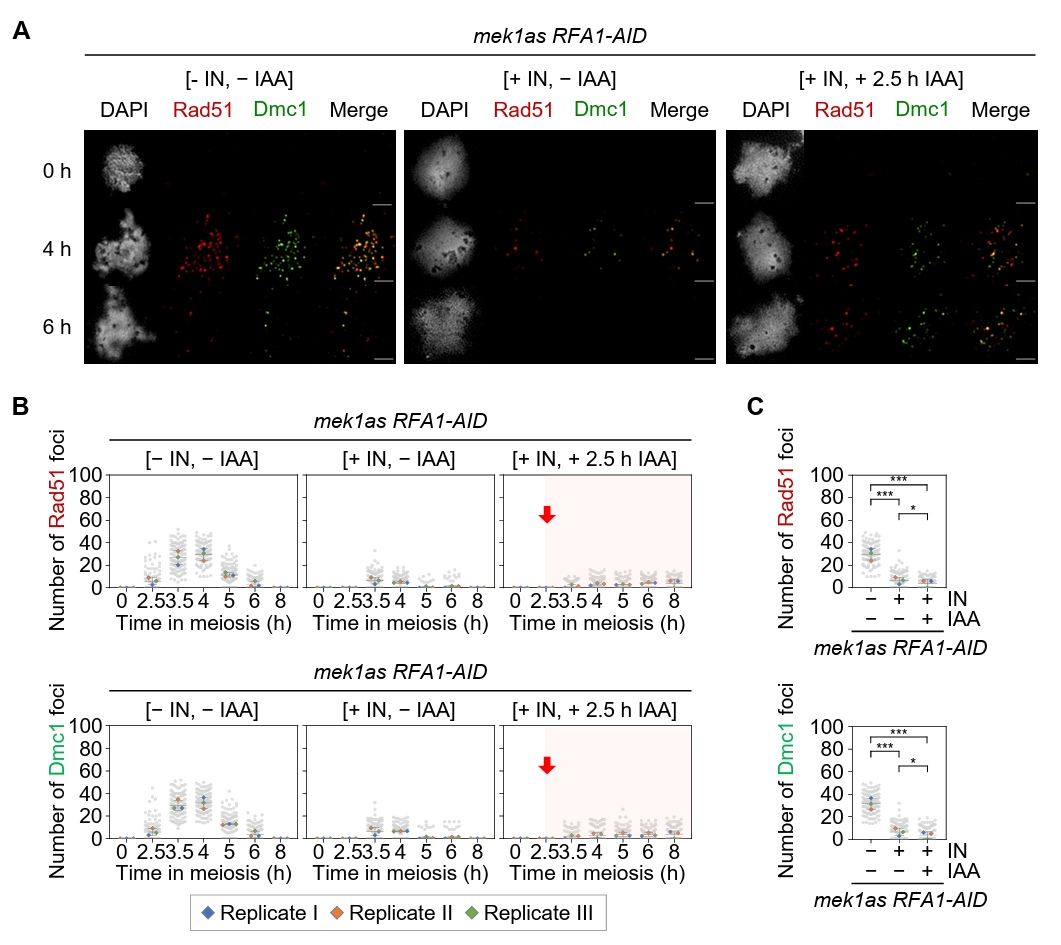
Fig. 5.Rfa1 is required for Rad51 and Dmc1 foci formation. (A) Representative images of meiotic chromosome spreads from the mek1as RFA1–AID strain, immuno-stained with anti-Rad51 (red) and anti-Dmc1 (green). Mek1 kinase activity was inhibited by the addition of 1-NA-PP1 (IN; 2 µM) at 0 h. CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to induce Rfa1 degradation. Scale bar = 2.5 µM. (B) Quantification of Rad51 and Dmc1 foci per nucleus in the mek1as RFA1–AID strain. Data are presented as the mean ± SD (three independent experiments). (C) Maximum peak time point of Rad51 and Dmc1 foci in the mek1as RFA1–AID strain shown in (B) [– IN, – IAA at 4 h], [+ IN, – IAA at 3.5 h] and [+ IN, + IAA at 8 h]. Data are presented as the mean ± SD (three independent experiments). P values for relevant comparisons are determined by Student’s t-test.
- Agarwal S, Roeder GS. 2000. Zip3 provides a link between recombination enzymes and synaptonemal complex proteins. Cell. 102: 245–255. ArticlePubMed
- Bishop AC, Buzko O, Shokat KM. 2001. Magic bullets for protein kinases. Trends Cell Biol. 11: 167–172. ArticlePubMed
- Bishop DK, Zickler D. 2004. Early decision; meiotic crossover interference prior to stable strand exchange and synapsis. Cell. 117: 9–15. ArticlePubMed
- Blat Y, Protacio RU, Hunter N, Kleckner N. 2002. Physical and functional interactions among basic chromosome organizational features govern early steps of meiotic chiasma formation. Cell. 111: 791–802. ArticlePubMed
- Borde V, de Massy B. 2013. Programmed induction of DNA double-strand breaks during meiosis: Setting up communication between DNA and the chromosome structure. Curr Opin Genet Dev. 23: 147–155. ArticlePubMed
- Börner GV, Kleckner N, Hunter N. 2004. Crossover/noncrossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of meiosis. Cell. 117: 29–45. ArticlePubMed
- Brill SJ, Stillman B. 1991. Replication factor-A from Saccharomyces cerevisiae is encoded by three essential genes coordinately expressed at S phase. Genes Dev. 5: 1589–1600. ArticlePubMed
- Busygina V, Sehorn MG, Shi IY, Tsubouchi H, Roeder GS, et al. 2008. Hed1 regulates Rad51-mediated recombination via a novel mechanism. Genes Dev. 22: 786–795. ArticlePubMedPMC
- Cannavo E, Cejka P, Kowalczykowski SC. 2013. Relationship of DNA degradation by Saccharomyces cerevisiae exonuclease 1 and its stimulation by RPA and Mre11-Rad50-Xrs2 to DNA end resection. Proc Natl Acad Sci USA. 110: E1661–E1668. ArticlePubMedPMC
- Cesar BI, Kim Y. 2026. Structure and function of the synaptonemal complex. J Cell Biol. 225: e202511222. ArticlePubMedPDF
- Chen SH, Albuquerque CP, Liang J, Suhandynata RT, Zhou H. 2010. A proteome-wide analysis of kinase-substrate network in the DNA damage response. J Biol Chem. 285: 12803–12812. ArticlePubMedPMC
- Chen H, Lisby M, Symington LS. 2013. RPA coordinates DNA end resection and prevents formation of DNA hairpins. Mol Cell. 50: 589–600. ArticlePubMedPMC
- Chen R, Wold MS. 2014. Replication protein A: Single-stranded DNA's first responder: Dynamic DNA-interactions allow replication protein A to direct single-strand DNA intermediates into different pathways for synthesis or repair. BioEssays. 36: 1156–1161. ArticlePubMedPMC
- Chen Z, Yang H, Pavletich NP. 2008. Mechanism of homologous recombination from the RecA-ssDNA/dsDNA structures. Nature. 453: 489–494. ArticlePubMedPDF
- Cho HR, Kong YJ, Hong SG, Kim KP. 2016. Hop2 and Sae3 are required for dmc1-mediated double-strand break repair via homolog bias during meiosis. Mol Cells. 39: 550–556. ArticlePubMedPMC
- Choi JE, Chung WH. 2026. Synthetic rescue in Saccharomyces cerevisiae: Concepts, large-scale genetic mapping, and functional implications. J Microbiol. 64: e2512017. ArticlePDF
- Chua PR, Roeder GS. 1998. Zip2, a meiosis-specific protein required for the initiation of chromosome synapsis. Cell. 93: 349–359. ArticlePubMed
- Conway AB, Lynch TW, Zhang Y, Fortin GS, Fung CW, et al. 2004. Crystal structure of a Rad51 filament. Nat Struct Mol Biol. 11: 791–796. ArticlePubMedPDF
- Crickard JB, Greene EC. 2018. Biochemical attributes of mitotic and meiotic presynaptic complexes. DNA Repair (Amst). 71: 148–157. ArticlePubMedPMC
- Crickard JB, Kaniecki K, Kwon Y, Sung P, Lisby M, et al. 2018. Regulation of Hed1 and Rad54 binding during maturation of the meiosis-specific presynaptic complex. EMBO J. 37: e98728. ArticlePubMedPMCLink
- Da Ines O, Bazile J, Gallego ME, White CI. 2022. DMC1 attenuates RAD51-mediated recombination in Arabidopsis. PLoS Genet. 18: e1010322. ArticlePubMedPMC
- De Muyt A, Jessop L, Kolar E, Sourirajan A, Chen J, et al. 2012. BLM helicase ortholog Sgs1 is a central regulator of meiotic recombination intermediate metabolism. Mol Cell. 46: 43–53. ArticlePubMedPMC
- Garcia V, Phelps SE, Gray S, Neale MJ. 2011. Bidirectional resection of DNA double-strand breaks by Mre11 and Exo1. Nature. 479: 241–244. ArticlePubMedPMCPDF
- Gold AL, Kim Y. 2026. Zipping up the synaptonemal complex: Pathways to homologous chromosome synapsis. BioEssays. 48: e70097. ArticlePubMedPMCLink
- Hinch AG, Becker PW, Li T, Moralli D, Zhang G, et al. 2020. The configuration of RPA, RAD51, and DMC1 binding in meiosis reveals the nature of critical recombination intermediates. Mol Cell. 79: 689–701. ArticlePubMedPMC
- Hollingsworth NM, Byers B. 1989. HOP1: A yeast meiotic pairing gene. Genetics. 121: 445–462. ArticlePubMedPMCPDF
- Hollingsworth NM, Gaglione R. 2019. The meiotic-specific Mek1 kinase in budding yeast regulates interhomolog recombination and coordinates meiotic progression with double-strand break repair. Curr Genet. 65: 631–641. ArticlePubMedPMCPDF
- Hong S, Joo JH, Yun H, Kim K. 2019a. The nature of meiotic chromosome dynamics and recombination in budding yeast. J Microbiol. 57: 221–231. ArticlePDF
- Hong S, Joo JH, Yun H, Kleckner N, Kim KP. 2019b. Recruitment of Rec8, Pds5 and Rad61/Wapl to meiotic homolog pairing, recombination, axis formation and S-phase. Nucleic Acids Res. 47: 11691–11708. ArticlePDF
- Hong S, Sung Y, Yu M, Lee M, Kleckner N, et al. 2013. The logic and mechanism of homologous recombination partner choice. Mol Cell. 51: 440–453. ArticlePubMedPMC
- Hunter N, Kleckner N. 2001. The single-end invasion: An asymmetric intermediate at the double-strand break to double-Holliday junction transition of meiotic recombination. Cell. 106: 59–70. ArticlePubMed
- Jessop L, Lichten M. 2008. Mus81/Mms4 endonuclease and Sgs1 helicase collaborate to ensure proper recombination intermediate metabolism during meiosis. Mol Cell. 31: 313–323. ArticlePubMedPMC
- Jo MK, Rhee K, Kim KP, Hong S. 2022. Yeast polyubiquitin unit regulates synaptonemal complex formation and recombination during meiosis. J Microbiol. 60: 705–714. ArticlePubMedPDF
- Joo JH, Hong S, Higashide MT, Choi EH, Yoon S, et al. 2024. RPA interacts with Rad52 to promote meiotic crossover and noncrossover recombination. Nucleic Acids Res. 52: 3794–3809. ArticlePubMedPMCPDF
- Joo JH, Kang HA, Kim KP, Hong S. 2022. Meiotic prophase roles of Pds5 in recombination and chromosome condensation in budding yeast. J Microbiol. 60: 177–186. ArticlePubMedPDF
- Joudeh L, Appleby RE, Maman JD, Pellegrini L. 2025. Structural mechanism of strand exchange by the RAD51 filament. eLife. 14: RP107114.ArticlePubMedPMCPDF
- Keeney S. 2001. Mechanism and control of meiotic recombination initiation. Curr Top Dev Biol. 52: 1–53. ArticlePubMed
- Keeney S, Lange J, Mohibullah N. 2014. Self-organization of meiotic recombination initiation: General principles and molecular pathways. Annu Rev Genet. 48: 187–214. ArticlePubMedPMC
- Kim KP, Weiner BM, Zhang L, Jordan A, Dekker J, et al. 2010. Sister cohesion and structural axis components mediate homolog bias of meiotic recombination. Cell. 143: 924–937. ArticlePubMedPMC
- Kong YJ, Joo JH, Kim KP, Hong S. 2017. Hed1 promotes meiotic crossover formation in Saccharomyces cerevisiae. J Microbiol Biotechnol. 27: 405–411. ArticlePubMed
- Kowalczykowski SC. 2015. An overview of the molecular mechanisms of recombinational DNA repair. Cold Spring Harb Perspect Biol. 7: a016410.ArticlePubMedPMC
- Lao JP, Cloud V, Huang CC, Grubb J, Thacker D, et al. 2013. Meiotic crossover control by concerted action of Rad51-Dmc1 in homolog template bias and robust homeostatic regulation. PLoS Genet. 9: e1003978. ArticlePubMedPMC
- Lao JP, Oh SD, Shinohara M, Shinohara A, Hunter N. 2008. Rad52 promotes postinvasion steps of meiotic double-strand-break repair. Mol Cell. 29: 517–524. ArticlePubMedPMC
- Latypov V, Rothenberg M, Lorenz A, Octobre G, Csutak O, et al. 2010. Roles of Hop1 and Mek1 in meiotic chromosome pairing and recombination partner choice in Schizosaccharomyces pombe. Mol Cell Biol. 30: 1570–1581. ArticlePubMedPMCLink
- Lee MS, Higashide MT, Choi H, Li K, Hong S, et al. 2021. The synaptonemal complex central region modulates crossover pathways and feedback control of meiotic double-strand break formation. Nucleic Acids Res. 49: 7537–7553. ArticlePubMedPMCPDF
- Lee MS, Joo JW, Choi H, Kang HA, Kim K. 2020. Mec1 modulates interhomolog crossover and interplays with Tel1 at post double-strand break stages. J Microbiol Biotechnol. 30: 469–475. ArticlePubMedPMC
- Lee MS, Joo JH, Kim K. 2017. Roles of budding yeast Hrr25 in recombination and sporulation. J Microbiol Biotechnol. 27: 1198–1203. ArticlePubMed
- Li X, Heyer WD. 2008. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res. 18: 99–113. ArticlePubMedPMCPDF
- Lord SJ, Velle KB, Mullins RD, Fritz-Laylin LK. 2020. SuperPlots: Communicating reproducibility and variability in cell biology. J Cell Biol. 219: e202001064. ArticlePubMedPMCPDF
- Machovina TS, Mainpal R, Daryabeigi A, McGovern O, Paouneskou D, et al. 2016. A surveillance system ensures crossover formation in C. elegans. Curr Biol. 26: 2873–2884. ArticlePubMedPMC
- Mimitou EP, Symington LS. 2008. Sae2, Exo1 and Sgs1 collaborate in DNA double-strand break processing. Nature. 455: 770–774. ArticlePubMedPMCPDF
- Moon HY, Sim GH, Kim HJ, Kim K, Kang HA. 2022. Assessment of Cre-lox and CRISPR-Cas9 as tools for recycling of multiple-integrated selection markers in Saccharomyces cerevisiae. J Microbiol. 60: 18–30. ArticlePubMedPDF
- Morawska M, Ulrich HD. 2013. An expanded tool kit for the auxin-inducible degron system in budding yeast. Yeast. 30: 341–351. ArticlePubMedPMCLink
- Morrical SW. 2015. DNA-pairing and annealing processes in homologous recombination and homology-directed repair. Cold Spring Harb Perspect Biol. 7: a016444.ArticlePubMedPMC
- Nadarajan S, Lambert TJ, Altendorfer E, Gao J, Blower MD, et al. 2017. Polo-like kinase-dependent phosphorylation of the synaptonemal complex protein SYP-4 regulates double-strand break formation through a negative feedback loop. eLife. 6: e23437. ArticlePubMedPMCPDF
- Neale MJ, Pan J, Keeney S. 2005. Endonucleolytic processing of covalent protein-linked DNA double-strand breaks. Nature. 436: 1053–1057. ArticlePubMedPMCPDF
- New JH, Sugiyama T, Zaitseva E, Kowalczykowski SC. 1998. Rad52 protein stimulates DNA strand exchange by Rad51 and replication protein A. Nature. 391: 407–410. ArticlePubMedPDF
- Nishimura K, Fukagawa T, Takisawa H, Kakimoto T, Kanemaki M. 2009. An auxin-based degron system for the rapid depletion of proteins in non-plant cells. Nat Methods. 6: 917–922. ArticlePubMedPDF
- Niu H, Li X, Job E, Park C, Moazed D, et al. 2007. Mek1 kinase is regulated to suppress double-strand break repair between sister chromatids during budding yeast meiosis. Mol Cell Biol. 27: 5456–5467. ArticlePubMedPMCLink
- Niu H, Wan L, Baumgartner B, Schaefer D, Loidl J, et al. 2005. Partner choice during meiosis is regulated by Hop1-promoted dimerization of Mek1. Mol Biol Cell. 16: 5804–5818. ArticlePubMedPMC
- Niu H, Wan L, Busygina V, Kwon Y, Allen JA, et al. 2009. Regulation of meiotic recombination via Mek1-mediated Rad54 phosphorylation. Mol Cell. 36: 393–404. ArticlePubMedPMC
- Oh SD, Lao JP, Hwang PY, Taylor AF, Smith GR, et al. 2007. BLM ortholog, Sgs1, prevents aberrant crossing-over by suppressing formation of multichromatid joint molecules. Cell. 130: 259–272. ArticlePubMedPMC
- Page SL, Hawley SR. 2004. The genetics and molecular biology of the synaptonemal complex. Annu Rev Cell Dev Biol. 20: 525–558. ArticlePubMed
- Pattabiraman D, Roelens B, Woglar A, Villeneuve AM. 2017. Meiotic recombination modulates the structure and dynamics of the synaptonemal complex during C. elegans meiosis. PLoS Genet. 13: e1006670. ArticlePubMedPMC
- Paull TT, Gellert M. 1998. The 3′ to 5′ exonuclease activity of Mre 11 facilitates repair of DNA double-strand breaks. Mol Cell. 1: 969–979. ArticlePubMed
- Rhee K, Choi H, Kim KP, Joo JH. 2023. A method for physical analysis of recombination intermediates in Saccharomyces cerevisiae. J Microbiol. 61: 939–951. ArticlePubMedPDF
- Rockmill B, Roeder GS. 1990. Meiosis in asynaptic yeast. Genetics. 126: 563–574. ArticlePubMedPMCPDF
- Sampathkumar A, Zhong C, Tang Y, Fujita Y, Ito M, et al. 2024. Replication protein-A, RPA, plays a pivotal role in the maintenance of recombination checkpoint in yeast meiosis. Sci Rep. 14: 9550.ArticlePubMedPMCPDF
- San Filippo J, Sung P, Klein H. 2008. Mechanism of eukaryotic homologous recombination. Annu Rev Biochem. 77: 229–257. ArticlePubMed
- Sanchez A, Reginato G, Cejka P. 2021. Crossover or non-crossover outcomes: Tailored processing of homologous recombination intermediates. Curr Opin Genet Dev. 71: 39–47. ArticlePubMed
- Sansam CL, Pezza RJ. 2015. Connecting by breaking and repairing: Mechanisms of DNA strand exchange in meiotic recombination. FEBS J. 282: 2444–2457. ArticlePubMedPMC
- Sheridan SD, Yu X, Roth R, Heuser JE, Sehorn MG, et al. 2008. A comparative analysis of Dmc1 and Rad51 nucleoprotein filaments. Nucleic Acids Res. 36: 4057–4066. ArticlePubMedPMC
- Shodhan A, Xaver M, Wheeler D, Lichten M. 2022. Turning coldspots into hotspots: Targeted recruitment of axis protein Hop1 stimulates meiotic recombination in Saccharomyces cerevisiae. Genetics. 222: iyac106.ArticlePubMedPMC
- Smith AV, Roeder GS. 1997. The yeast red1 protein localizes to the cores of meiotic chromosomes. J Cell Biol. 136: 957–967. ArticlePubMedPMCPDF
- Sung P. 1994. Catalysis of ATP-dependent homologous DNA pairing and strand exchange by yeast RAD51 protein. Science. 265: 1241–1243. ArticlePubMed
- Sung P. 1997. Function of yeast Rad52 protein as a mediator between replication protein A and the Rad51 recombinase. J Biol Chem. 272: 28194–28197. ArticlePubMed
- Sym M, Engebrecht JA, Roeder GS. 1993. ZIP1 is a synaptonemal complex protein required for meiotic chromosome synapsis. Cell. 72: 365–378. ArticlePubMed
- Sym M, Roeder GS. 1995. Zip1-induced changes in synaptonemal complex structure and polycomplex assembly. J Cell Biol. 128: 455–466. ArticlePubMedPMCPDF
- Symington LS, Rothstein R, Lisby M. 2014. Mechanisms and regulation of mitotic recombination in Saccharomyces cerevisiae. Genetics. 198: 795–835. ArticlePubMedPMCPDF
- Tamai T, Reginato G, Ojiri R, Morita I, Avrutis A, et al. 2024. Sae2 controls Mre11 endo- and exonuclease activities by different mechanisms. Nat Commun. 15: 7221.ArticlePubMedPMCPDF
- Terentyev Y, Johnson R, Neale MJ, Khisroon M, Bishop-Bailey A, et al. 2010. Evidence that MEK1 positively promotes interhomologue double-strand break repair. Nucleic Acids Res. 38: 4348–4360. Article
- Voelkel-Meiman K, Cheng SY, Parziale M, Morehouse SJ, Feil A, et al. 2019. Crossover recombination and synapsis are linked by adjacent regions within the N terminus of the Zip1 synaptonemal complex protein. PLoS Genet. 15: e1008201. ArticlePubMedPMC
- Wan L, de los Santos T, Zhang C, Shokat K, Hollingsworth NM. 2004. Mek1 kinase activity functions downstream of RED1 in the regulation of meiotic double strand break repair in budding yeast. Mol Biol Cell. 15: 11–23. ArticlePubMed
- Wang W, Daley JM, Kwon Y, Krasner DS, Sung P. 2017. Plasticity of the Mre11-Rad50-Xrs2-Sae2 nuclease ensemble in the processing of DNA-bound obstacles. Genes Dev. 31: 2331–2336. ArticlePubMed
- Wold MS. 1997. Replication protein A: A heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu Rev Biochem. 66: 61–92. ArticlePubMed
- Xiao W, Chow BL, Rathgeber L. 1996. The repair of DNA methylation damage in Saccharomyces cerevisiae. Curr Genet. 30: 461–468. ArticlePubMedPDF
- Yoon SW, Lee MS, Xaver M, Zhang L, Hong SG, et al. 2016. Meiotic prophase roles of Rec8 in crossover recombination and chromosome structure. Nucleic Acids Res. 44: 9296–9314. ArticlePubMedPMC
- Zakharyevich K, Ma Y, Tang S, Hwang PY, Boiteux S, et al. 2010. Temporally and biochemically distinct activities of Exo1 during meiosis: Double-strand break resection and resolution of double Holliday junctions. Mol Cell. 40: 1001–1015. ArticlePubMedPMC
- Zhu Z, Chung WH, Shim EY, Lee SE, Ira G. 2008. Sgs1 helicase and two nucleases Dna2 and Exo1 resect DNA double-strand break ends. Cell. 134: 981–994. ArticlePubMedPMC
- Zickler D, Kleckner N. 1999. Meiotic chromosomes: integrating structure and function. Annu Rev Genet. 33: 603–754. ArticlePubMed
- Zickler D, Kleckner N. 2015. Recombination, pairing, and synapsis of homologs during meiosis. Cold Spring Harb Perspect Biol. 7: a016626.ArticlePubMedPMC
- Zimmerman ES, Schulman BA, Zheng N. 2010. Structural assembly of cullin-RING ubiquitin ligase complexes. Curr Opin Struct Biol. 20: 714–721. ArticlePubMedPMC
References
Supplementary Information
References
Citations
Citations to this article as recorded by 
 ePub Link
ePub Link Cite this Article
Cite this Article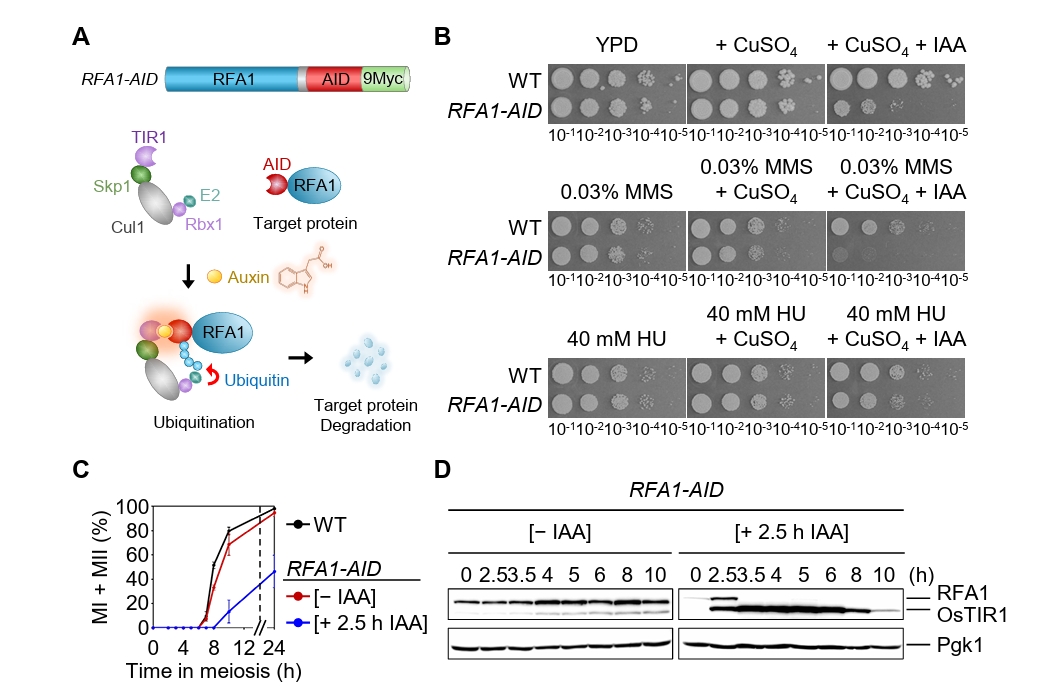
Prophase roles of replication protein A in crossover formation and meiotic progression
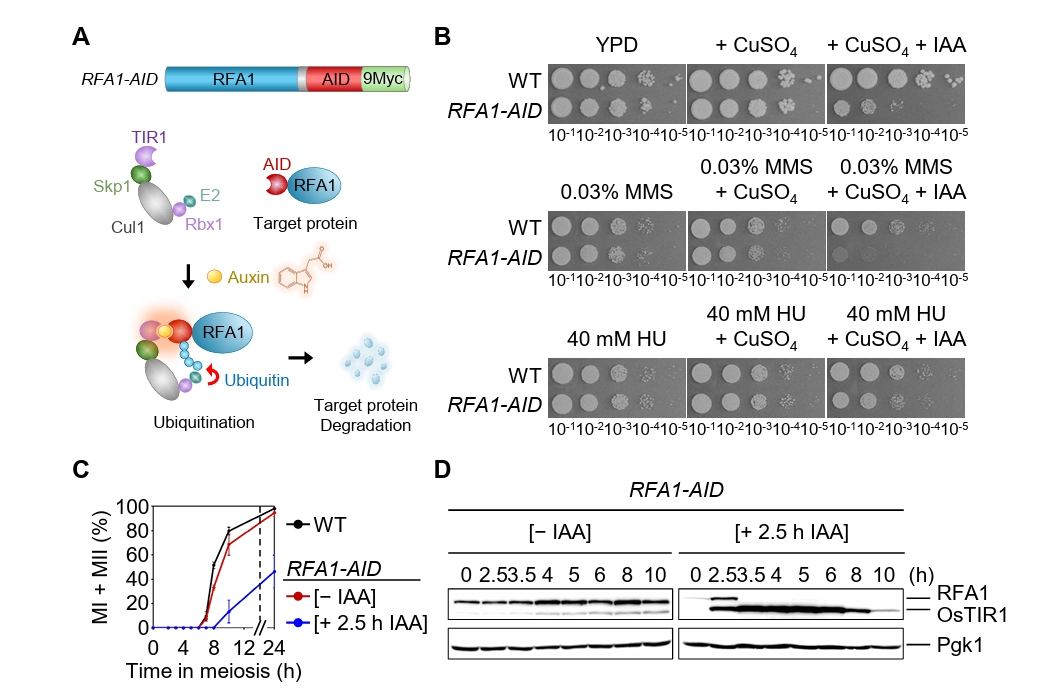
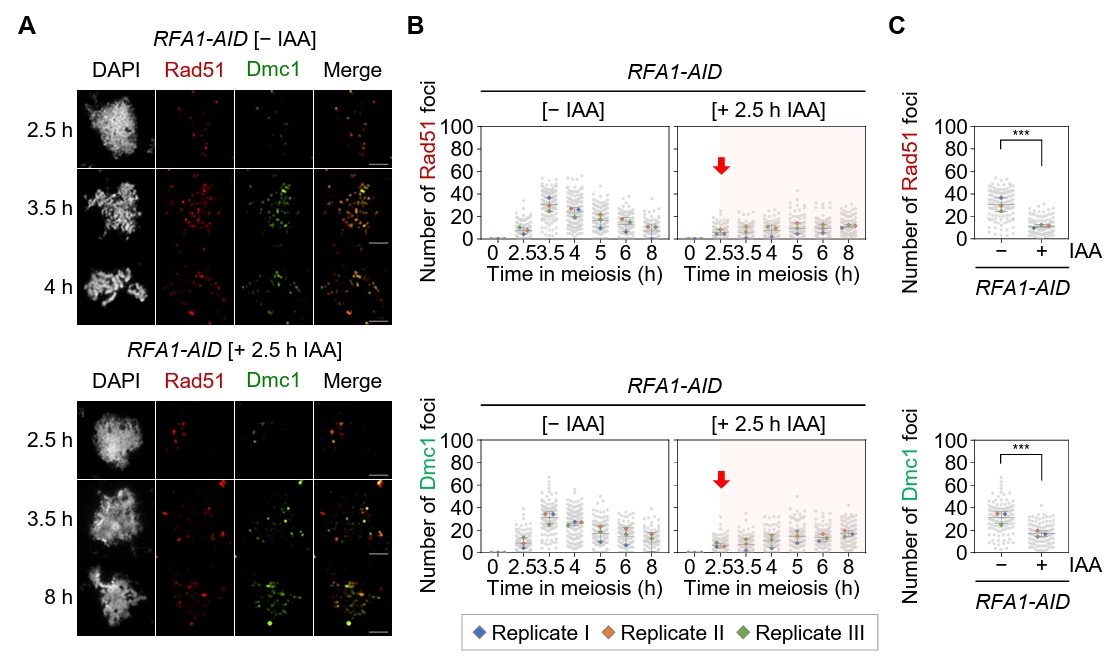
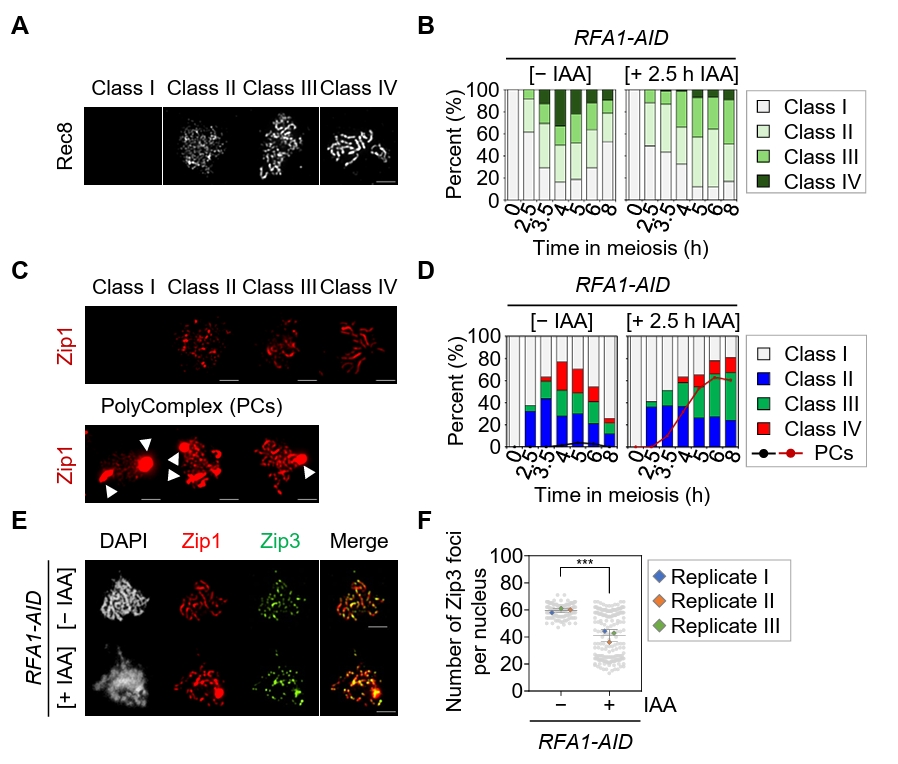
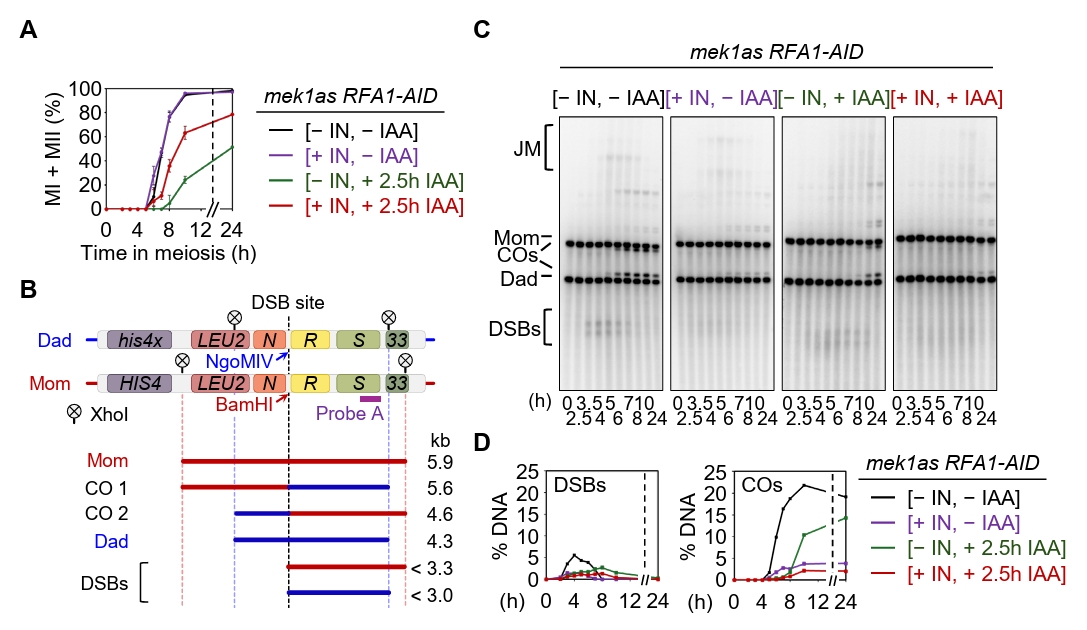
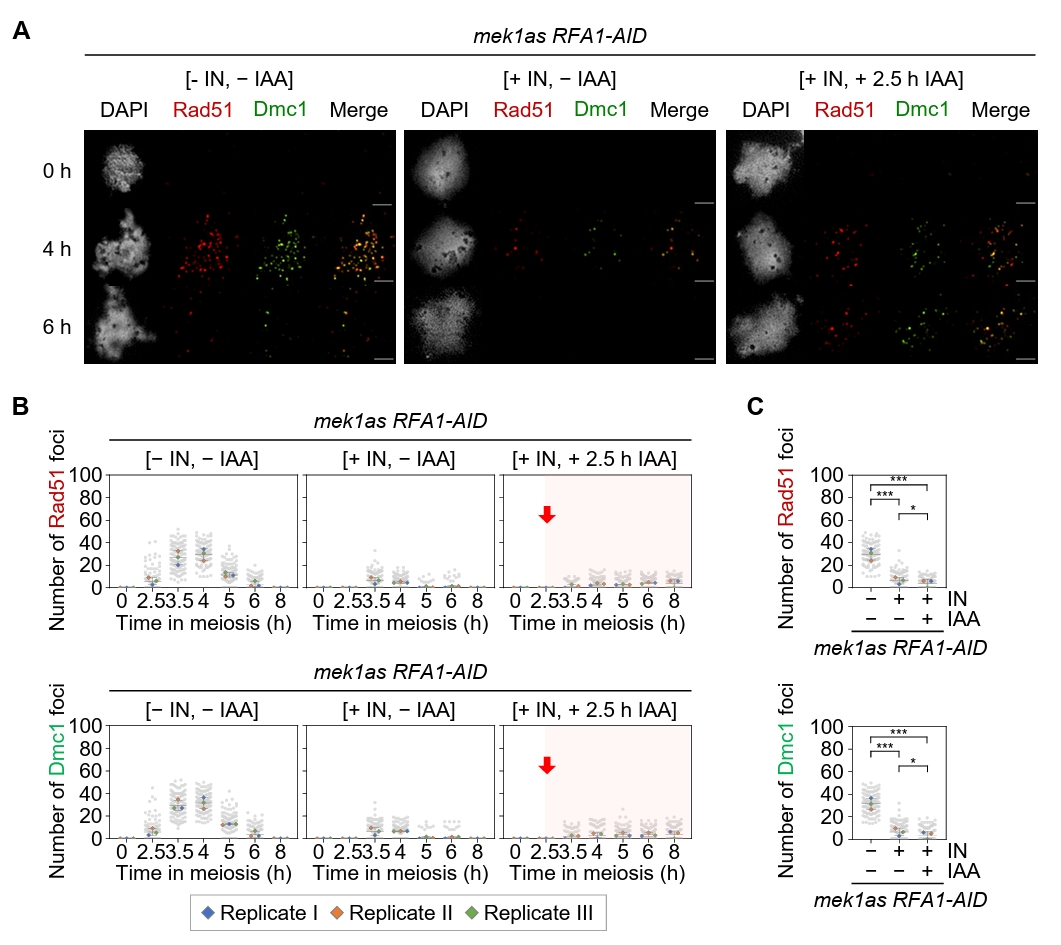
Fig. 1. Conditional Rfa1 degradation via auxin-inducible degron alters DNA damage sensitivity and meiotic progression. (A) Schematic representation of the RFA1–AID construct and the auxin-inducible degron (AID) system. Upon auxin (IAA) treatment, RFA1–AID is ubiquitinated by the SCFTIR1 complex and subsequently degraded by the proteasome. (B) DNA damage sensitivity assay of wild-type (WT) and RFA1–AID strains. The 10-fold serial dilutions (from 10-1 to 10-5) were spotted onto YPD plates containing the DNA-damaging agents: 0.03% MMS, 40 mM HU, 20 μM CuSO4, or 2 mM IAA. Plates were incubated at 30°C for 2 days. (C) Meiotic nuclear divisions in WT and RFA1–AID strains. Data are presented as the mean ± standard deviation (SD) (N > 200/time point; three independent biological replicates). (D) Protein expression levels of Rfa1 and Pgk1 during meiosis in the RFA1–AID strain. CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, followed by IAA (2 mM) treatment at 2.5 h to induce Rfa1 degradation. Proteins were detected using anti-Myc (for RFA1–AID–9Myc) and anti-Pgk1 antibodies. Unprocessed blot images are shown in Fig. S1B.
Fig. 2. Rfa1 is required for the formation of Rad51 and Dmc1 foci during meiotic recombination. (A) Representative immunofluorescence image of a meiotic nuclear spread from RFA1–AID strains in the absence [– IAA] or presence [+ IAA] of IAA. Cells were stained with anti-Rad51 (red) and anti-Dmc1 (green). CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to promote Rfa1 degradation. Scale bar = 2.5 μm. (B) Quantification of Rad51 and Dmc1 foci per nucleus in the RFA1–AID strain during meiotic recombination. Data are presented as the mean ± SD (three independent experiments). (C) Maximum peak time point of Rad51 and Dmc1 foci in the RFA1–AID strain shown in (B) [– IAA at 3.5 h] and [+ IAA at 8 h]. Data are presented as the mean ± SD (three independent experiments). P values for relevant comparisons are determined by Student’s t-test.
Fig. 3. Depletion of Rfa1 causes defects in meiotic recombination progression. (A) Representative immunofluorescence images of meiotic chromosome spreads stained with anti-Rec8 in the RFA1–AID strain. CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to induce Rfa1 degradation. Nuclear spreads were classified into four categories based on the Rec8 staining pattern: Class I, no staining; Class II, a modest number of chromosomes; Class III, extended or short linear chromosomes; Class IV, a full array of chromosomes. Scale bar = 2.5 μm. (B) Quantification of Rec8 staining classes at each time point shown in (A). (C) Representative images of meiotic chromosome spreads stained with anti-Zip1 in the RFA1–AID strain. CuSO4 and IAA were added as in (A). Nuclear spreads were classified into four categories based on the Zip1 staining pattern: Class I, blank; Class II, dotty chromosomes; Class III, short or discontinuous linear chromosomes; Class IV, full-array chromosomes. White arrowheads indicate polycomplexes (PCs). Scale bar = 2.5 μm. (D) Quantification of Zip1 staining classes at each time point shown in (C). The line graph indicates the percentage of cells with PCs. (E) Representative meiotic chromosome spreads from RFA1–AID strains immunostained with anti-Zip1 (red) and anti-Flag (for Zip3 staining, green). CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to induce Rfa1 degradation. Scale bar = 2.5 μm. (F) Quantification of Zip3 foci per nucleus at the peak time points of Zip3 expression during meiotic recombination: [– IAA] at 4 h and [+ IAA] at 6 h. Data are presented as the mean ± SD (three independent experiments). P values for relevant comparisons are determined by Student’s t-test.
Fig. 4. Rfa1 is essential for meiotic DSB and crossover formation. (A) Meiotic nuclear division in the mek1as RFA1–AID strain. Mek1 kinase activity was inhibited by the addition of 1-NA-PP1 (IN; 2 µM) at 0 h. CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to induce degradation of Rfa1. Data are presented as the mean ± SD (N > 200 cells per time point; three independent biological replicates). (B) Schematic of the HIS4::LEU2 recombination hotspot, which generates double-strand breaks (DSBs) during meiosis. Positions of XhoI restriction sites are indicated by an X. Probe A (purple), used for Southern blot analysis, is shown. DSB, double-strand break; CO, crossover. (C) Representative one-dimensional gel images from mek1as RFA1–AID strains. (D) Quantification of DSB and CO levels shown in (C).
Fig. 5. Rfa1 is required for Rad51 and Dmc1 foci formation. (A) Representative images of meiotic chromosome spreads from the mek1as RFA1–AID strain, immuno-stained with anti-Rad51 (red) and anti-Dmc1 (green). Mek1 kinase activity was inhibited by the addition of 1-NA-PP1 (IN; 2 µM) at 0 h. CuSO4 (20 μM) was added at 2 h to induce OsTIR1 expression, and IAA (2 mM) was added at 2.5 h to induce Rfa1 degradation. Scale bar = 2.5 µM. (B) Quantification of Rad51 and Dmc1 foci per nucleus in the mek1as RFA1–AID strain. Data are presented as the mean ± SD (three independent experiments). (C) Maximum peak time point of Rad51 and Dmc1 foci in the mek1as RFA1–AID strain shown in (B) [– IN, – IAA at 4 h], [+ IN, – IAA at 3.5 h] and [+ IN, + IAA at 8 h]. Data are presented as the mean ± SD (three independent experiments). P values for relevant comparisons are determined by Student’s t-test.
Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Prophase roles of replication protein A in crossover formation and meiotic progression
TOP

